ņä£ļĪĀ
ņØśļŻīĻĖ░ņłĀņØś ļ░£ļŗ¼Ļ│╝ ĒĢ©Ļ╗ś ĻĖ░ļīĆņŚ¼ļ¬ģņØ┤ ļ╣Āļź┤Ļ▓ī ņ”ØĻ░ĆĒĢśļ®┤ņä£ ņĀä ņäĖĻ│äņĀüņØĖ Ļ│ĀļĀ╣ĒÖöĻ░Ć Ļ░ĆņåŹĒÖöļÉśĻ│Ā ņ׳ļŗż[1,2]. ņØ┤ļĪ£ ņØĖĒĢ┤ ļŗżņ¢æĒĢ£ ņŗĀĻ▓ĮĻ│ä ņ¦łĒÖśļōżņØś ņ£Āļ│æļźĀņØ┤ ĻĖēĻ▓®ĒĢśĻ▓ī ņ”ØĻ░ĆĒĢśĻ│Ā ņ׳ļŖöļŹ░, ĒŖ╣Ē׳ ĒīīĒé©ņŖ©ļ│æ(Parkinson disease)Ļ│╝ ņĢīņĖĀĒĢśņØ┤ļ©Ėļ│æ(Alzheimer disease) ļō▒ņØś ņŗĀĻ▓ĮĒć┤Ē¢ēņ¦łĒÖśņØś ņ”ØĻ░ĆņäĖĻ░Ć Ļ░ĆĒīīļź┤ļŗż[3]. ņŗĀĻ▓ĮĒć┤Ē¢ēņ¦łĒÖśņØś Ļ│ĄĒåĄņĀüņØĖ ņ£äĒŚśņØĖņ×ÉĻ░Ć ļ░öļĪ£ ļéśņØ┤ņØ┤ĻĖ░ ļĢīļ¼ĖņØ┤ļŗż.
Ļ│ĀļĀ╣ņŚÉņä£ ņŻ╝ļĪ£ ļ░£ņāØĒĢśļŖö ņŗĀĻ▓ĮĒć┤Ē¢ēņ¦łĒÖśļōżņØĆ ņ£ĀņĀäņĀüņØĖ Ļ░Éņłśņä▒(genetic susceptibility)Ļ│╝ ĒÖśĻ▓ĮņĀü ņÜöņØĖ(environmental factor)ņØś ņāüĒśĖņ×æņÜ®ņ£╝ļĪ£ ļ░£ņāØĒĢśĻĖ░ ļĢīļ¼ĖņŚÉ ņØ╝ļČĆ ņ£ĀņĀä ņ¦łĒÖśņØä ņĀ£ņÖĖĒĢśĻ│ĀļŖö ļīĆļČĆļČä ņøÉņØĖņØä ĒĢśļéśļĪ£ ĒŖ╣ņĀĢĒĢśĻĖ░ ņ¢┤ļĀĄļŗż. ņĢīĒīīņŗ£ļłäĒü┤ļĀłņØĖ(╬▒-synuclein), ĒāĆņÜ░ļŗ©ļ░▒(tau protein) ļō▒ņØ┤ ņŗĀĻ▓ĮĒć┤Ē¢ēņ¦łĒÖśņØś ņŻ╝ņÜö ĻĖ░ņ░®ņ¦ĆļØ╝ļŖö Ļ▓āņØ┤ ļ░ØĒśĆņĪīņ¦Ćļ¦ī, ņØ┤ļōżņØ┤ ņ¢┤ļööņä£ļČĆĒä░ ņāØĻ▓©ļéśņä£ ņ¢┤ļ¢╗Ļ▓ī ļ│æņØä ļ¦īļōżņ¢┤Ļ░ĆļŖöņ¦ĆņŚÉ ļīĆĒĢ£ Ļ│╝ņĀĢņŚÉ ļīĆĒĢ┤ņä£ļŖö ņĢäņ¦ü ļČłļ¬ģĒÖĢĒĢ£ ļČĆļČäļōżņØ┤ ļ¦Äļŗż.
ņןņØĆ ļ¬Ė ņåŹņŚÉ ņ׳ņ¦Ćļ¦ī ļÅÖņŗ£ņŚÉ Ēö╝ļČĆņ▓śļ¤╝ ņÖĖļČĆ ĒÖśĻ▓ĮĻ│╝ ņĀæĒĢśĻ│Ā ņ׳ļŗż. ņŻ╝ļĪ£ ņØīņŗØļ¼╝ņØä ņåīĒÖöņŗ£ņ╝£ ņśüņ¢æļČäņØä ĒØĪņłśĒĢśļŖö ņŚŁĒĢĀņØä ĒĢśĻĖ░ ļĢīļ¼ĖņŚÉ ļ¦żņÜ░ ļ¦ÄņØĆ ņØ╝ņØä ĒĢśļŖö ĻĖ░Ļ┤ĆņØ┤ļŗż. ļŗżņ¢æĒĢ£ ņäĖĻĘĀļōżĻ│╝ ĒĢ©Ļ╗ś Ļ│ĄņĪ┤ĒĢ┤ņĢ╝ ĒĢśļŖö ĒÖśĻ▓ĮņØ┤ļ»ĆļĪ£ ņØ┤ļōżĻ│╝ ĒĢäņŚ░ņĀüņ£╝ļĪ£ ņśüĒ¢źņØä ņŻ╝Ļ│Āļ░øņØä ņłśļ░¢ņŚÉ ņŚåļŗż. ņŗØņŖĄĻ┤ĆņØ┤ ņןļé┤ņäĖĻĘĀņ┤ØņØś ĻĄ¼ņä▒ņŚÉ ņśüĒ¢źņØä ņŻ╝Ļ│Ā, ņןļé┤ņäĖĻĘĀņ┤ØņØĆ ņןņÜ┤ļÅÖ, ņØīņŗØļ¼╝ ļ░Å ņĢĮļ¼╝ņØś ĒØĪņłś ļō▒ņŚÉ Ļ┤ĆņŚ¼ĒĢ£ļŗż.
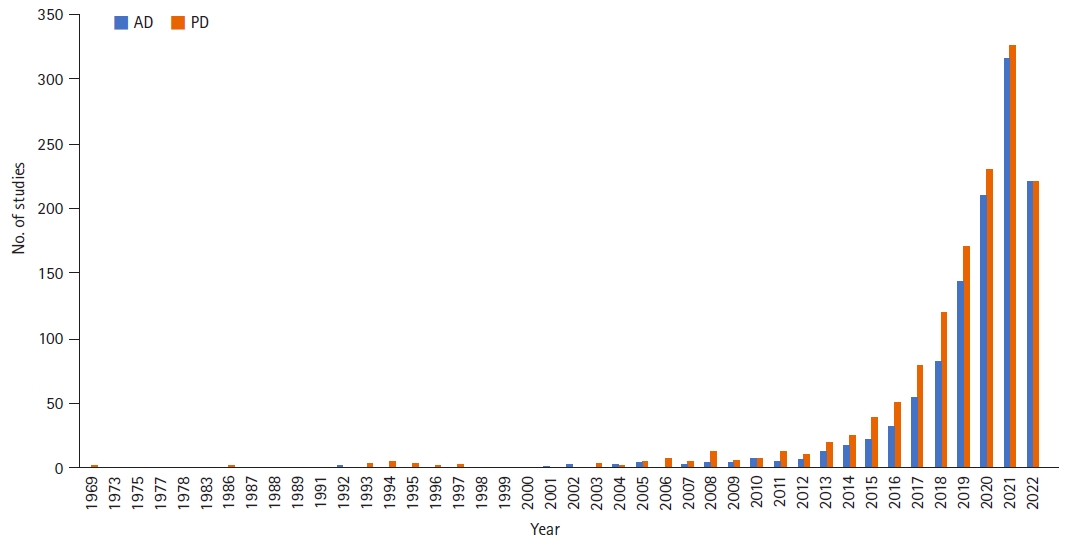
ņä£ļĪ£ ļ®Ćļ”¼ ļ¢©ņ¢┤ņĀĖ ņ׳ļŖö Ļ▓āņ▓śļ¤╝ ļ│┤ņśĆļŹś ņŗĀĻ▓ĮĻ│äņÖĆ ņןņØ┤ ņĄ£ĻĘ╝ ņŚ¼ļ¤¼ ņŚ░ĻĄ¼ļź╝ ĒåĄĒĢ┤ ņä£ļĪ£ ĻĖ┤ļ░ĆĒĢśĻ▓ī ņśüĒ¢źņØä ņŻ╝Ļ│Āļ░øļŖöļŗżļŖö Ļ▓āņØ┤ ņĢīļĀżņ¦ĆĻ▓ī ļÉśņŚłļŗż. PubMed (https://pubmed.ncbi.nlm.nih.gov/)ņŚÉņä£ 2022ļģä 8ņøö 6ņØ╝ Ēśäņ×¼ ŌĆśgut & AlzheimerŌĆÖņÖĆ ŌĆśgut & ParkinsonŌĆÖņ£╝ļĪ£ Ļ▓ĆņāēĒĢ£ Ļ▓░Ļ│╝ Ļ░üĻ░ü 1,041Ļ▒┤, 1,232Ļ▒┤ņØś ņŚ░ĻĄ¼Ļ░Ć Ļ▓ĆņāēļÉśņŚłĻ│Ā 2010ļģä ņØ┤ĒøäņŚÉ ĒÅŁļ░£ņĀüņ£╝ļĪ£ ņŚ░ĻĄ¼Ļ░Ć ņ”ØĻ░ĆĒĢśņśĆņØīņØä ņĢī ņłś ņ׳ļŗż(Figure 1). ĒŖ╣Ē׳ ņŗĀĻ▓ĮĒć┤Ē¢ēņ¦łĒÖśņØś ļ░£ņāØņŚÉ ņ׳ņ¢┤ ņןĻ│╝ ņןļé┤ņäĖĻĘĀņ┤ØņØś ņŚŁĒĢĀņØä Ļ░ĢņĪ░ĒĢśļŖö ņŚ░ĻĄ¼ļōżņØ┤ ļ│┤Ļ│ĀļÉśļ®┤ņä£ Ļ┤Ćņŗ¼ņØ┤ ļŹöĒĢ┤ņ¦ĆĻ│Ā ņ׳ļŖö ņāüĒÖ®ņØ┤ļŗż. ļ│Ė ņóģņäżņŚÉņä£ļŖö ļīĆĒæ£ņĀüņØĖ ņŗĀĻ▓ĮĒć┤Ē¢ēņ¦łĒÖśņØĖ ĒīīĒé©ņŖ©ļ│æĻ│╝ ņĢīņĖĀĒĢśņØ┤ļ©Ėļ│æņØś ļ░£ļ│æņŚÉ ņןņØ┤ ņ¢┤ļ¢ż ņśüĒ¢źņØä ņŻ╝ļŖöņ¦ĆņŚÉ ļīĆĒĢ£ ļé┤ņÜ®ņØä ņĀĢļ”¼ĒĢ┤ ļ│┤Ļ│Āņ×É ĒĢ£ļŗż.
ņןņ£╝ļĪ£ļČĆĒä░ ņŗĀĻ▓ĮĒć┤Ē¢ēņ¦łĒÖśņØś ļ│æļ”¼ļ│ĆĒÖöĻ░Ć ņŗ£ņ×æĒĢśļŖöĻ░Ć?
1. ĒīīĒé©ņŖ©ļ│æ
1) ņĀäĻĄ¼ņ”Øņāü
ĒīīĒé©ņŖ©ļ│æņØĆ ņŻ╝ļĪ£ ņĢīĒīīņŗ£ļłäĒü┤ļĀłņØĖĻ│╝ ļŻ©ņØ┤ņåīņ▓┤(Lewy body)Ļ░Ć ņŗĀĻ▓ĮņäĖĒżņŚÉ ņ╣©ņ░®ļÉśĻ│Ā, ņØ┤ļĪ£ ņØĖĒĢ┤ ĒĢ┤ļŗ╣ ņŗĀĻ▓ĮņäĖĒżļōżņØ┤ ņé¼ļ®ĖĒĢśļ®┤ņä£ ļéśĒāĆļéśļŖö ņ¦łĒÖśņØ┤ļŗż[4]. ņØ╝ļ░śņĀüņ£╝ļĪ£ ĒÖśņ×ÉĻ░Ć ņŗĀĻ▓ĮĻ│╝ ņØśņé¼ļź╝ ņ░ŠņĢäņśżļŖö ņŗ£ĻĖ░ļŖö ļŖÉļ”¼Ļ│Ā(bradykinesia), ļ¢©ļ”¼Ļ│Ā(tremor at rest), ļ╗Żļ╗ŻĒĢśĻ│Ā(rigidity), ĻĄ¼ļČĆņĀĢĒĢ£ ņ×ÉņäĖ(stooped posture)ļź╝ ņĘ©ĒĢśĻ▓ī ļÉĀ ļĢī(ņ”ē ņÜ┤ļÅÖņ”ØņāüņØ┤ ļéśĒāĆļéĀ ļĢī)ņØ┤ņ¦Ćļ¦ī, ļ¦ÄņØĆ ĒÖśņ×ÉļōżņØ┤ ņÜ┤ļÅÖņ”ØņāüņØ┤ ļéśĒāĆļéśĻĖ░ ņØ┤ņĀäļČĆĒä░ ļ│Ćļ╣ä(constipation), ĒøäĻ░üņåīņŗż(hyposmia), ņÜ░ņÜĖņ”Ø(depression), ļĀśņłśļ®┤ņןņĢĀ(rapid eye movement sleep behavior disorder) ļō▒ņØś ņĀäĻĄ¼ņ”ØņāüņØä Ļ▓¬ļŖö Ļ▓āņ£╝ļĪ£ ņĢīļĀżņĀĖ ņ׳ļŗż[5,6].
ņØ┤ļ¤¼ĒĢ£ ņĀäĻĄ¼ņ”Øņāü ņżæņŚÉ Ļ░Ćņן ņØ┤ļźĖ ņŗ£ĻĖ░ņŚÉ ļéśĒāĆļéśļŖö ņ”ØņāüņØ┤ ļ│Ćļ╣äņØ┤ļŗż. ņŚ░ĻĄ¼ļ¦łļŗż ņ░©ņØ┤ļŖö ņ׳ņ£╝ļéś ļīĆļץ ņÜ┤ļÅÖņ”ØņāüņØ┤ ļéśĒāĆļéśĻĖ░ 15ļģä ņØ┤ņāü ņĀäļČĆĒä░ ļéśĒāĆļéśļŖö Ļ▓āņ£╝ļĪ£ ļ│┤Ļ│ĀļÉ£ļŗż[6]. Braak stage IņŚÉņä£ļŖö ļ»ĖņŻ╝ņŗĀĻ▓ĮņØś ļō▒ņ¬ĮņÜ┤ļÅÖņŗĀĻ▓ĮĒĢĄ(dorsal motor nucleus of the vagus)ņØ┤ ņ╣©ļ▓öļÉśļ»ĆļĪ£ ļ│Ćļ╣äĻ░Ć ņ£Āļ░£ļÉ£ļŗżĻ│Ā ļ│╝ ņłśļÅä ņ׳Ļ▓Āņ¦Ćļ¦ī[7], ĻĘĖ ņØ┤ĒĢś ļĀłļ▓©ņØĖ ļ»ĖņŻ╝ņŗĀĻ▓Į(vagus nerve)ņØ┤ļéś ņןņŗĀĻ▓Įņ┤Ø(enteric plexus)ņØ┤ ņ╣©ļ▓öļÉśņ¢┤ļÅä ļ│Ćļ╣äļŖö ļ░£ņāØĒĢĀ ņłś ņ׳ļŗż.
2) ņןņ£╝ļĪ£ļČĆĒä░ ļćīĻ╣īņ¦Ć
ĒīīĒé©ņŖ©ļ│æņŚÉņä£ ļŻ©ņØ┤ņåīņ▓┤Ļ░Ć ņןļé┤ņŚÉņä£ļÅä ļ░£Ļ▓¼ļÉśņŚłļŗżļŖö ņŚ░ĻĄ¼ļŖö ņĢĮ 40ļģä ņĀäļČĆĒä░ ņ׳ņŚłļŗż[8,9]. ņŗØļÅäļČĆĒä░ ņ¦üņןĻ╣īņ¦Ć Ļ▒░ņØś ņĀä ņ£äņןĻ┤ĆņŚÉ Ļ▒Ėņ│É ļŻ©ņØ┤ņåīņ▓┤Ļ░Ć ļ░£Ļ▓¼ļÉśņŚłņ£╝ļéś, ĒĢ£ļÅÖņĢł ņĀüĻĘ╣ņĀüņØĖ ĒøäņåŹ ņŚ░ĻĄ¼ļŖö ņ¦äĒ¢ēļÉśņ¦Ć ņĢŖņĢśļŗż. 2006ļģä Braak ļō▒[10]ņØ┤ ĒīīĒé©ņŖ©ļ│æ ĒÖśņ×ÉņŚÉņä£ ņĀÉļ¦ēĒĢśņŗĀĻ▓Įņ┤Ø(submucosal plexus)Ļ│╝ ĻĘ╝ņ£ĪņĖĄņŗĀĻ▓Įņ┤Ø(myenteric plexus)ņŚÉ ļŻ©ņØ┤ņåīņ▓┤Ļ░Ć ņ╣©ņ░®ļÉśņŚłņØīņØä ļŗżņŗ£ ĒĢ£ļ▓ł ļ│┤Ļ│ĀĒĢśņśĆĻ│Ā, ņØ┤ ļŻ©ņØ┤ņåīņ▓┤Ļ░Ć ļ»ĖņŻ╝ņŗĀĻ▓ĮņØä ĒåĄĒĢ┤ ņżæņČöņŗĀĻ▓ĮĻ│äļĪ£ ņĀäļŗ¼ļÉśņŚłņØä Ļ░ĆļŖźņä▒ņØä ņĀ£ĻĖ░ĒĢśņśĆļŗż. ņ”ē ņÖĖļČĆ ĒÖśĻ▓ĮĻ│╝ ļ¦×ļŗ┐ņĢä ņ׳ļŖö ņןņ£╝ļĪ£ļČĆĒä░ ņĢīĒīīņŗ£ļłäĒü┤ļĀłņØĖĻ│╝ ļŻ©ņØ┤ņåīņ▓┤ņØś ņ╣©ņ░®ņØ┤ ņŗ£ņ×æļÉśĻ│Ā ņżæņČöņŗĀĻ▓ĮĻ│äļĪ£ ņØ┤ņ¢┤ņ¦ĆļŖö ĒåĄļĪ£ņØĖ ļ»ĖņŻ╝ņŗĀĻ▓ĮņØä ĒåĄĒĢ┤ ņĀäļŗ¼ļÉśņŚłņØä Ļ▓āņØ┤ļØ╝ļŖö Ļ░ĆņäżņØ┤ļŗż.
ņØ┤ Ļ░ĆņäżņØ┤ ņ×ģņ”ØļÉśĻĖ░ ņ£äĒĢ┤ņä£ļŖö ļ¬ć Ļ░Ćņ¦Ć ņ”Øļ¬ģņØ┤ ĒĢäņÜöĒĢśļŗż. ņÜ░ņäĀ, ĒīīĒé©ņŖ©ļ│æ ĒÖśņ×ÉņØś ņ£äņןĻ┤ĆņŚÉņä£ ĒīīĒé©ņŖ©ļ│æņØ┤ ņŚåļŖö ĻĘĖļŻ╣ņŚÉ ļ╣äĒĢ┤ ņĢīĒīīņŗ£ļłäĒü┤ļĀłņØĖĻ│╝ ļŻ©ņØ┤ņåīņ▓┤ ņ╣©ņ░®ņØ┤ ĒÖĢņŚ░ĒĢ┤ņĢ╝ ĒĢ£ļŗż. ņŚ░ĻĄ¼ļ¦łļŗż ļŗżņåī ņ░©ņØ┤ļŖö ņ׳ņ£╝ļéś Braak ļō▒[10]ņØś ļ│┤Ļ│Ā ņØ┤Ēøä ņ¦äĒ¢ēļÉ£ ļīĆļČĆļČäņØś ņŚ░ĻĄ¼ņŚÉņä£ ĒīīĒé©ņŖ©ļ│æņŚÉņä£ņØś ņ£äņןĻ┤Ć ļ│æļ”¼ļ│ĆĒÖöĻ░Ć ļæÉļō£ļ¤¼ņĪīļŗż[11]. ļæśņ¦Ė, ņĢīĒīīņŗ£ļłäĒü┤ļĀłņØĖ ņ╣©ņ░®ņ£╝ļĪ£ ņØĖĒĢ£ ļ│æļ”¼ļ│ĆĒÖöĻ░Ć ļ»ĖņŻ╝ņŗĀĻ▓ĮņØä ĒåĄĒĢ┤ ņĀäļŗ¼ļÉĀ ņłś ņ׳ņ¢┤ņĢ╝ ĒĢ£ļŗż. ņźÉņÖĆ ņøÉņłŁņØ┤ļź╝ ņØ┤ņÜ®ĒĢ£ ļÅÖļ¼╝ņŗżĒŚśņØä ĒåĄĒĢ┤ ņ£äņןĻ┤ĆņŚÉ ļŻ©ņØ┤ņåīņ▓┤ ņ£Āņé¼ņ▓┤ļź╝ ņŻ╝ņé¼ĒĢśņśĆņØä ļĢī ļ»ĖņŻ╝ņŗĀĻ▓ĮņØä ĒåĄĒĢ┤ ļćīļĪ£ ņĀäļŗ¼ļÉ©ņØä ĒÖĢņØĖĒĢ£ ņŚ░ĻĄ¼ļōżņØ┤ ņ׳ļŗż[12,13]. ņģŗņ¦Ė, ļ»ĖņŻ╝ņŗĀĻ▓ĮņØä ņĀłļŗ©ĒĢśļ®┤ ļ│æļ”¼ļ│ĆĒÖöĻ░Ć ņĀäļŗ¼ļÉśņ¦Ć ņĢŖņĢäņĢ╝ ĒĢ£ļŗż. ņźÉļź╝ ņØ┤ņÜ®ĒĢ£ ņŗżĒŚśņŚÉņä£ ņ£äņןĻ┤ĆņŚÉ ņĢīĒīīņŗ£ļłäĒü┤ļĀłņØĖņØä ņŻ╝ņ×ģĒĢ£ ņØ┤Ēøä ļ»ĖņŻ╝ņŗĀĻ▓ĮņØä ņĀłļŗ©ĒĢ£ Ļ▓ĮņÜ░ ņÖĖņØĖņä▒ ņĢīĒīīņŗ£ļłäĒü┤ļĀłņØĖņŚÉ ņØśĒĢ£ ļ│æļ”¼ļ│ĆĒÖöĻ░Ć ļćīņŚÉņä£ ļ░£ņāØĒĢśņ¦Ć ņĢŖņØīņØä ĒÖĢņØĖĒĢśņśĆļŗż[14]. ņé¼ļ×īņŚÉĻ▓īņä£ ļÅÖņØ╝ĒĢ£ ņŗżĒŚśņØä ĒĢĀ ņłśļŖö ņŚåņ¦Ćļ¦ī, ļ»ĖņŻ╝ņŗĀĻ▓ĮņĀłņĀ£ņłĀ(vagotomy)ņØä ļ░øņØĆ ņé¼ļ×īļōżņØä ņłśļģäĻ░ä ņČöņĀüĻ┤Ćņ░░ĒĢ£ Ļ▓░Ļ│╝ ņżäĻĖ░ļ»ĖņŻ╝ņŗĀĻ▓ĮņĀłņĀ£ņłĀ(truncal vagotomy)ņØä ņŗ£Ē¢ēĒĢ£ ĻĘĖļŻ╣ņŚÉņä£ ĒīīĒé©ņŖ©ļ│æņØś ļ░£ņāØļźĀņØ┤ ļé«ņØīņØä ĒÖĢņØĖĒĢśņŚ¼, Ļ░äņĀæņĀüņ£╝ļĪ£ ĒīīĒé©ņŖ©ļ│æ ļ│æļ”¼ļ│ĆĒÖöņØś ņŗ£ņ×æņØ┤ ņ£äņןĻ┤ĆņØ╝ Ļ░ĆļŖźņä▒ņØä ņĀ£ņŗ£ĒĢśņśĆļŗż[15,16].
3) Brain-first versus body-first
ĒīīĒé©ņŖ©ļ│æņØś ņĀäĻĄ¼ņ”Øņāü ņżæņŚÉ Ļ░Ćņן ĒīīĒé©ņŖ©ļ│æņŚÉ ĒŖ╣ņØ┤ņĀüņØĖ ņ”ØņāüņØĆ ļĀśņłśļ®┤ņןņĢĀņØ┤ļŗż. ļĀśņłśļ®┤ņןņĢĀĻ░Ć ņ׳ļŖö Ļ▓ĮņÜ░ņØś ņāüļīĆņ£äĒŚśļÅäĻ░Ć 50 ņĀĢļÅäļĪ£ ļ¬©ļōĀ ņĀäĻĄ¼ņ”Øņāü ņżæņŚÉ Ļ░Ćņן ļåÆļŗż[6]. ļö░ļØ╝ņä£ ļĀśņłśļ®┤ņןņĢĀĻ░Ć ņ׳ņ¦Ćļ¦ī ņĢäņ¦ü ĒīīĒé©ņŖ©ļ│æņØś ņÜ┤ļÅÖņ”ØņāüņØ┤ ļéśĒāĆļéśņ¦Ć ņĢŖņØĆ ņāüĒÖ®ņŚÉņä£ ņŗĀņ▓┤ņØś ņ¢┤ļŖÉ ļČĆņ£äļź╝ ļŹö ļ¦ÄņØ┤ ņ╣©ļ▓öĒ¢łļŖöņ¦Ćļź╝ ņĢīļ®┤ ļ│æņØś ņ¦äĒ¢ē ļ░®Ē¢źņØä Ļ░ĆļŖĀĒĢĀ ņłś ņ׳ņØä Ļ▓āņØ┤ļŗż.
2019ļģä ļ│Ćļ╣ä, ļĀśņłśļ®┤ņןņĢĀ, ņ×Éņ£©ņŗĀĻ▓ĮĻ│äņ”ØņāüņØä Ļ░Ćņ¦ä ņĀäĻĄ¼ļŗ©Ļ│äņØś ļŻ©ņØ┤ņåīņ▓┤ņŚ░Ļ┤Ćņ¦łĒÖś ņØśņŗ¼ĒÖśņ×Éļź╝ ļīĆņāüņ£╝ļĪ£ ļÅäĒīīļ»╝ņÜ┤ļ░śņ▓┤ņśüņāü(dopamine transporter scan)Ļ│╝ metaiodobenzylguanidine ņŗ¼ĻĘ╝ņŖżņ║ö(MIBG myocardial cintigraphy)ņØä ņŗ£Ē¢ēĒĢ£ ņŚ░ĻĄ¼Ļ░Ć ņ׳ļŗż[17]. ĒĢ┤ļŗ╣ ņŚ░ĻĄ¼ņŚÉņä£ ņĀäņ▓┤ ļīĆņāüĒÖśņ×É 18ļ¬ģ ņżæņŚÉ ļÅäĒīīļ»╝ņÜ┤ļ░śņ▓┤ņśüņāüņŚÉņä£ ņ¢æņä▒ņØä ļ│┤ņØĖ Ļ▓ĮņÜ░ļŖö 56% (10ļ¬ģ)ņØĖ ļ░śļ®┤ņŚÉ, MIBGņŗ¼ĻĘ╝ņŖżņ║öņŚÉņä£ ņ¢æņä▒ņØä ļ│┤ņØĖ Ļ▓ĮņÜ░ļŖö 94% (17ļ¬ģ)ņØ┤ņŚłļŗż. ĒĢ┤ļČĆĒĢÖņĀü ņ£äņ╣śļź╝ Ļ│ĀļĀżĒĢśļ®┤ ņÜ┤ļÅÖņ”ØņāüņØä ļīĆļ│ĆĒĢśļŖö ĒØæņāēņ¦łņØś ļÅäĒīīļ»╝ ņäĖĒżņØś ņé¼ļ®ĖņØ┤ ļ│ĖĻ▓®ņĀüņ£╝ļĪ£ ņØ╝ņ¢┤ļéśĻĖ░ ņĀäņŚÉ ņØ┤ļ»Ė ņŗ¼ņןņØś ņ×Éņ£©ņŗĀĻ▓ĮņØ┤ ņśüĒ¢źņØä ļ░øņĢśļŗżļŖö ļ£╗ņØ┤ļ»ĆļĪ£ ļ│æļ”¼ļ│ĆĒÖöĻ░Ć ļ¬Ėņ£╝ļĪ£ļČĆĒä░ ņČ£ļ░£ĒĢśņŚ¼ ļćīļĪ£ ņ¦äĒ¢ēĒĢśņśĆļŗżļŖö Ļ░ĆņäżņØ┤ ĒלņØä ņ¢╗ļŖöļŗż(body-first type).
ĻĘĖļ¤¼ļéś ļ¬©ļōĀ ĒīīĒé©ņŖ©ļ│æ ĒÖśņ×ÉļōżņØ┤ ļĀśņłśļ®┤ņןņĢĀļź╝ Ļ░Ćņ¦ĆĻ│Ā ņ׳ļŖö Ļ▓āņØĆ ņĢäļŗłļŗż. ņĢīĒīīņŗ£ļłäĒü┤ļĀłņØĖ ļ│æļ”¼ļ│ĆĒÖöĻ░Ć ņŻ╝ļ│Ćņ£╝ļĪ£ ļ▓łņĀĖ ļéśĻ░ĆļŖö ĒŖ╣ņä▒ņØä Ļ░ÉņĢłĒĢĀ ļĢī, ļĀśņłśļ®┤ņןņĢĀĻ░Ć ņŚåļŖö ĒīīĒé©ņŖ©ļ│æ ĒÖśņ×ÉļōżņØĆ ļĀśņłśļ®┤ņןņĢĀĻ░Ć ņ׳ļŖö Ļ▓ĮņÜ░ņÖĆ ļ│æļ”¼ņØś ņČ£ļ░£ņ¦ĆņĀÉņØ┤ ļŗżļź┤ļŗżĻ│Ā ņāØĻ░üĒĢĀ ņłś ņ׳ļŗż. ņØ┤ļ¤¼ĒĢ£ Ļ░ĆņäżņØä ļ░öĒāĢņ£╝ļĪ£ ĒŖ╣ļ░£ņä▒ļĀśņłśļ®┤ņןņĢĀņŚÉ ļĀśņłśļ®┤ņןņĢĀĻ░Ć ļÅÖļ░śļÉ£ ĒīīĒé©ņŖ©ļ│æĻ│╝ ĻĘĖļĀćņ¦Ć ņĢŖņØĆ ĒīīĒé©ņŖ©ļ│æņØä ņČöĻ░ĆĒĢśņŚ¼ MIBGņŗ¼ĻĘ╝ņŖżņ║öĻ│╝ donepezil positron emission tomography/computed tomography (PET/CT)ļź╝ ņłśĒ¢ēĒĢ£ ņŚ░ĻĄ¼Ļ░Ć ņ׳ļŗż[18,19]. MIBGņŗ¼ĻĘ╝ņŖżņ║öņØĆ ņśłņāüļīĆļĪ£ ĒīīĒé©ņŖ©ļ│æ ņŚ¼ļČĆņÖĆ Ļ┤ĆĻ│äņŚåņØ┤ ļĀśņłśļ®┤ņןņĢĀĻ░Ć ņ׳ņ£╝ļ®┤ ļīĆļČĆļČä ņØ┤ņāüņØ┤ ļéśĒāĆļé¼Ļ│Ā, ļĀśņłśļ®┤ņןņĢĀĻ░Ć ņŚåļŖö ĒīīĒé©ņŖ©ļ│æņŚÉņä£ļŖö ņāüļīĆņĀüņ£╝ļĪ£ ņĀĢņāüņØĖ Ļ▓ĮņÜ░Ļ░Ć ļ¦ÄņĢśļŗż. ņןņÜ┤ļÅÖņØä ļ░śņśüĒĢśļŖö donepezil PET/CTļÅä MIBGņŗ¼ĻĘ╝ņŖżņ║ö Ļ▓░Ļ│╝ņÖĆ ņ£Āņé¼ĒĢ£ Ēī©Ēä┤ņØä ļ│┤ņśĆļŗż. Ļ▓░ĻĄŁ ļĀśņłśļ®┤ņןņĢĀĻ░Ć ņŚåļŖö ĒīīĒé©ņŖ©ļ│æņØĆ ņŗĀņ▓┤ņØś ņĢäļל ļČĆļČäņ£╝ļĪ£ļČĆĒä░ ļ│æļ”¼Ļ░Ć ņ¦äĒ¢ēļÉśņ¢┤ ņś©ļŗżļŖö ĻĘ╝Ļ▒░Ļ░Ć ļČĆņĪ▒ĒĢśļŗż(brain-first type).
ļ│æļ”¼ļ│ĆĒÖöņØś ņŗ£ņ×æņ¦ĆņĀÉņØä ņżæņŗ¼ņ£╝ļĪ£ ĒīīĒé©ņŖ©ļ│æņØś ĒāĆņ×ģņØä ĻĄ¼ļČäĒĢśļŖö ļ░®ņŗØņØ┤ ļ░öļĪ£ brain-first versus body-first Ļ░ĆņäżņØ┤ļŗż. ņØ┤ļź╝ ļÆĘļ░øņ╣©ĒĢśļŖö ļŗżņ¢æĒĢ£ ņŗżĒŚśņĀü, ņ×äņāüņĀü ĻĘ╝Ļ▒░ļōżņØ┤ ņ׳ņ£╝ļéś ļ╣äĒīÉņĀüņØĖ ņŗ£Ļ░üļÅä ņĪ┤ņ×¼ĒĢ£ļŗż[20]. ļ╣äļĪØ ļĀśņłśļ®┤ņןņĢĀĻ░Ć ļŻ©ņØ┤ņåīņ▓┤ņŚ░Ļ┤Ćņ¦łĒÖśņŚÉ ĒŖ╣ņØ┤ņĀüņØ┤ļØ╝Ļ│ĀļŖö ĒĢśņ¦Ćļ¦ī ņĢäļ░ĆļĪ£ņØ┤ļō£(amyloid)ļéś ĒāĆņÜ░ļŗ©ļ░▒ Ļ┤ĆļĀ© ņ¦łĒÖśņŚÉņä£ļÅä ļ│┤Ļ│ĀļÉ£ ļ░ö ņ׳ņ¢┤ ĒŚłņĀÉņØ┤ ņĪ┤ņ×¼ĒĢ£ļŗż. ņØ┤ļ│┤ļŗż ļŹö ņżæņÜöĒĢ£ Ļ▓āņØĆ ļĀśņłśļ®┤ņןņĢĀĻ░Ć ņ׳ļŖö ĒÖśņ×ÉņØś ļćīņŚÉ ņØ┤ļ»Ė ņĢīĒīīņŗ£ļłäĒü┤ļĀłņØĖ ņ╣©ņ░®Ļ│╝ ņŚ░Ļ┤ĆļÉ£ ļ│æļ”¼ļ│ĆĒÖöņÖĆ ņäĖĒżņé¼ļ®ĖņØ┤ ļ░£Ļ▓¼ļÉśņŚłļŗżļŖö ņĀÉņØ┤ļŗż. Ļ▓░ĻĄŁ ņØ┤ņ¦łņä▒(heterogeneity)ņØä Ļ░Ćņ¦ä ĒīīĒé©ņŖ©ļ│æņØ┤ļØ╝ļŖö Ēü░ ņ¦łĒÖśĻĄ░ņØä ļŗ©ņł£ĒĢ£ ļ░®ņŗØņ£╝ļĪ£ ļéśļłäļŖö Ļ▓ā ņ░©ņ▓┤Ļ░Ć ņ¢┤ļĀĄļŗżļŖö Ļ▓āņØ┤ļŗż.
2. ņĢīņĖĀĒĢśņØ┤ļ©Ėļ│æ
ĒīīĒé©ņŖ©ļ│æĻ│╝ ļŗ¼ļ”¼ ņĢīņĖĀĒĢśņØ┤ļ©Ėļ│æņØś ļ│æļ”¼ļ│ĆĒÖöĻ░Ć ņןņ£╝ļĪ£ļČĆĒä░ ņŗ£ņ×æĒĢśļŖöņ¦Ć ņŚ¼ļČĆļŖö ļČłļČäļ¬ģĒĢśļŗż[21]. ņØ╝ļČĆ ņןļé┤ņŚÉņä£ ĒāĆņÜ░ļŗ©ļ░▒ņØ┤ ļ░£Ļ▓¼ļÉśņŚłļŗżļŖö ļ│┤Ļ│ĀĻ░Ć ņ׳ĻĖ░ļŖö ĒĢśņ¦Ćļ¦ī[22,23], ļŗżļźĖ ņŚ░ĻĄ¼ņŚÉņä£ļŖö ņĢīņĖĀĒĢśņØ┤ļ©Ėņ╣śļ¦żņÖĆ ļ╣äņĢīņĖĀĒĢśņØ┤ļ©Ėņ╣śļ¦ż ĻĄ░ ļ¬©ļæÉņŚÉņä£ ļ░£Ļ▓¼ļÉśņ¦Ć ņĢŖņĢäņä£ ņØ╝Ļ┤ĆļÉ£ Ļ▓░Ļ│╝ļŖö ņĢäļŗłļŗż[24]. ņĄ£ĻĘ╝ ņŗ£Ē¢ēĒĢ£ ņŚ░ĻĄ¼ņŚÉņä£ļŖö ņןļé┤ ĒāĆņÜ░ļŗ©ļ░▒ņØ┤ ĒāĆņÜ░Ļ┤ĆļĀ© ņŗĀĻ▓ĮĒć┤Ē¢ēņ¦łĒÖśĻ│╝ ļīĆņĪ░ĻĄ░ ļ¬©ļæÉņŚÉņä£ ļ░£Ļ▓¼ļÉśņŚłņ£╝ļéś, ņ¢æ ĻĄ░ ņé¼ņØ┤ņØś ņ░©ņØ┤ļŖö ņŚåņŚłĻĖ░ ļĢīļ¼ĖņŚÉ ņ¦łĒÖśĒŖ╣ņØ┤ņä▒ņØĆ ņĀüņØĆ Ļ▓āņ£╝ļĪ£ ņāØĻ░üļÉ£ļŗż[25]. ļŹöĻĄ¼ļéś ĒīīĒé©ņŖ©ļ│æņŚÉņä£ņÖĆļŖö ļŗ¼ļ”¼ ņżäĻĖ░ļ»ĖņŻ╝ņŗĀĻ▓ĮņĀłņĀ£ņłĀņØä ņŗ£Ē¢ēĒĢ£ ĻĘĖļŻ╣ņŚÉņä£ ņĢīņĖĀĒĢśņØ┤ļ©Ėļ│æņØś ļ░£ņāØ ĒÖĢļźĀņØ┤ ļŹö ļåÆņ¦Ć ņĢŖņĢśĻĖ░ ļĢīļ¼ĖņŚÉ ņé¼ļ×ī ļīĆņāüņØś ņ×äņāüņŚ░ĻĄ¼ņŚÉņä£ļÅä ĻĘ╝Ļ▒░ļÅä ļČĆņĪ▒ĒĢśļŗż[26]. ĒīīĒé©ņŖ©ļ│æņŚÉņä£ļŖö ļ│Ćļ╣äĻ░Ć ņÜ┤ļÅÖņ”ØņāüņØä ņäĀĒ¢ēĒĢśļŖö ļ░śļ®┤ ņĢīņĖĀĒĢśņØ┤ļ©Ėļ│æņŚÉņä£ļŖö ņØ┤ļ¤¼ĒĢ£ Ļ┤ĆĻ│äĻ░Ć ļ¬ģĒÖĢņ╣ś ņĢŖņØĆ ņĀÉļÅä ņØ┤ļź╝ ļÆĘļ░øņ╣©ĒĢ£ļŗż.
ņŗĀĻ▓ĮĒć┤Ē¢ēņ¦łĒÖśņØś ļ░£ņāØņŚÉ ņśüĒ¢źņØä ņŻ╝ļŖö ĻĖ░ĒāĆ ņןņØś ņŚŁĒĢĀ
ĒīīĒé©ņŖ©ļ│æņŚÉņä£ ņןņØś ļ│æļ”¼ļ│ĆĒÖöĻ░Ć ļ»ĖņŻ╝ņŗĀĻ▓ĮņØä ĒåĄĒĢ┤ ļćīļĪ£ ņĀäļŗ¼ļÉśļŖö Ļ▓āņØ┤ ņŻ╝ņÜö ņøÉņØĖņØ╝ ņłś ņ׳ļŗżļŖö Ļ░ĆņäżņØ┤ ļ¦żņÜ░ ļ¦żļĀźņĀüņØ┤ņ¢┤ņä£ ļłłĻĖĖņØ┤ Ļ░Ćņ¦Ćļ¦ī ņןņØś ņŚŁĒĢĀņØ┤ ļŗ©ņ¦Ć ļ│æļ”¼ļ│ĆĒÖöņØś ņČ£ļ░£ņĀÉņŚÉļ¦ī ļ®łņČ░ ņ׳ļŖö Ļ▓āņØĆ ņĢäļŗłļŗż. ņĢīņĖĀĒĢśņØ┤ļ©Ėļ│æņØä ļ╣äļĪ»ĒĢ£ ļŗżļźĖ ņŗĀĻ▓ĮĒć┤Ē¢ēņ¦łĒÖśņŚÉņä£ļÅä ņן Ēś╣ņØĆ ņןļé┤ ļ¦łņØ┤Ēü¼ļĪ£ļ░öņØ┤ņś┤(gut microbiome)ņØś ļ│ĆĒÖöĻ░Ć ļ│æņØś ļ░£ņāØĻ│╝ ņ¦äĒ¢ēņŚÉ ņśüĒ¢źņØä ņżĆļŗżļŖö ņŚ░ĻĄ¼Ļ▓░Ļ│╝ļōżņØä ļ│╝ ļĢī, ļ»ĖņŻ╝ņŗĀĻ▓ĮņØä ĒåĄĒĢ£ ĻĖ░ņĀä ņÖĖņŚÉļÅä ņŗĀĻ▓ĮĒć┤Ē¢ēņ¦łĒÖśņŚÉ ņśüĒ¢źņØä ņŻ╝ļŖö ļŗżņ¢æĒĢ£ Ļ░äņĀæ Ļ▓ĮļĪ£Ļ░Ć ņĪ┤ņ×¼ĒĢĀ Ļ▓āņØ┤ļØ╝ļŖö ņĀÉņØä ņ¦Éņ×æĒĢĀ ņłś ņ׳ļŗż[27].
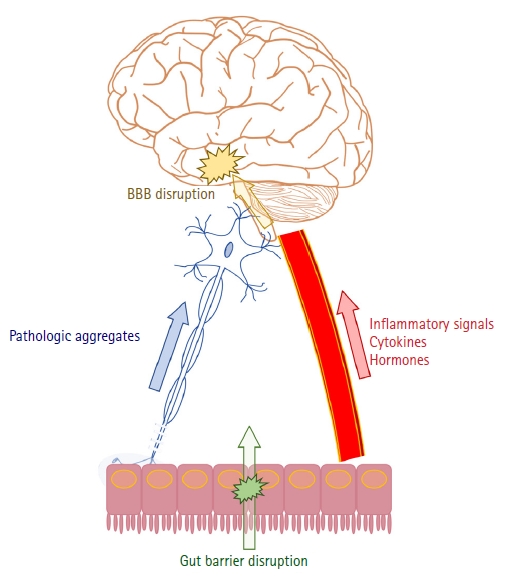
ĻĘĖ ņżæ Ļ░Ćņן ņżæņÜöĒĢ£ ļČĆļČäņØä ņ░©ņ¦ĆĒĢśļŖö Ļ▓āņØĆ ņŚ╝ņ”ØņØ┤ļŗż. ņĀäņŗĀ Ēś╣ņØĆ ņŗĀĻ▓ĮĻ│ä ņŚ╝ņ”ØņØ┤ ĒīīĒé©ņŖ©ļ│æĻ│╝ ņĢīņĖĀĒĢśņØ┤ņ¢┤ļ│æņØä ļ╣äļĪ»ĒĢ£ ļŗżņ¢æĒĢ£ ņŗĀĻ▓ĮĒć┤Ē¢ēņ¦łĒÖśņØś ļ░£ņāØņŚÉ ņśüĒ¢źņØä ņżĆļŗżļŖö Ļ▓āņØ┤ ņל ņĢīļĀżņĀĖ ņ׳ļŗż[28]. ņןņØ┤ ņØ┤ļ¤¼ĒĢ£ ņŚ╝ņ”Ø ļ│ĆĒÖöņŚÉ ņ׳ņ¢┤ ņżæņÜöĒĢ£ ņŚŁĒĢĀņØä ļŗ┤ļŗ╣ĒĢ£ļŗżļŖö Ļ▓āņØ┤ļŗż. ņןļé┤ ļ¦łņØ┤Ēü¼ļĪ£ļ░öņØ┤ņś┤ņØĆ Ļ░üņóģ ļ¦żĻ░£ļ¼╝ņ¦łņØä ĒåĄĒĢśņŚ¼ ņłÖņŻ╝ņØś ļ®┤ņŚŁĻĖ░ļŖźņŚÉ ņśüĒ¢źņØä ņżĆļŗż[29]. ņśłļź╝ ļōżņ¢┤ ņןļé┤ ņäĖĻĘĀņŚÉņä£ ļČäļ╣äļÉśļŖö ņ¦Ćļŗżļŗ╣(lipopolysaccharide)ņØĆ Ēśłļźśļź╝ Ļ▒░ņ│É ļćīņŚÉ ņŗĀĻ▓ĮņŚ╝ņ”ØņØä ņ£Āļ░£ĒĢĀ ņłś ņ׳ļŗż[30]. ņäĖĻĘĀņØś ņ¦Ćļŗżļŗ╣ņ£╝ļĪ£ ņØĖĒĢ┤ ĒśłņĢĪļćīņןļ▓Į(blood-brain barrier)ņØś Ēł¼Ļ│╝ļÅäĻ░Ć ņ”ØĻ░ĆĒĢśņŚ¼ ņŚ╝ņ”ØņäĖĒżņØś ņ¦äņ×ģņØ┤ ņÜ®ņØ┤ĒĢ┤ņ¦ĆĻĖ░ ļĢīļ¼ĖņØ┤ļŗż[31]. ņןļé┤ ņäĖĻĘĀņ£╝ļĪ£ļČĆĒä░ ĻĖ░ņØĖĒĢ£ ļŗ©ņćäņ¦Ćļ░®ņé░(short-chain fatty acid)ļÅä ļćīņŚÉņä£ņØś ļ®┤ņŚŁĻĖ░ļŖźņØä ļŗ┤ļŗ╣ĒĢśļŖö ļ»ĖņäĖĻĄÉņäĖĒż(microglia)ņØś ņä▒ņłÖņŚÉ Ļ┤ĆņŚ¼ĒĢ©ņ£╝ļĪ£ņŹ© ņŚ╝ņ”Ø ņĪ░ņĀłņŚÉ ņśüĒ¢źņØä ņżĆļŗż. ņØ┤ņ▓śļ¤╝ ņןĻ│╝ Ļ┤ĆļĀ©ļÉ£ ņŗĀĻ▓ĮĻ│ä ņŚ╝ņ”ØņØ┤ ņĢīņĖĀĒĢśņØ┤ļ©Ėļ│æ ļ░£ņāØņŚÉ ļÜ£ļĀĘĒĢ£ ņśüĒ¢źņØä ņżĆļŗżļŖö Ļ▓āņØĆ ņŚ¼ļ¤¼ ņŚ░ĻĄ¼ļź╝ ĒåĄĒĢ┤ ļ░ØĒśĆņĪīņ£╝ļ®░[32], ĒĢŁņāØņĀ£ ņ╣śļŻīļéś Ļ▒┤Ļ░ĢĒĢ£ ņןņØś ļ¦łņØ┤Ēü¼ļĪ£ļ░öņØ┤ņś┤ ņØ┤ņŗØņØ┤ ņŚ╝ņ”ØņØś Ļ░ÉņåīņÖĆ ļ│æļ”¼ļ│ĆĒÖöļź╝ ņżäņŚ¼ņŻ╝ņŚłļŗżļŖö ļÅÖļ¼╝ņŗżĒŚś Ļ▓░Ļ│╝ļź╝ ĒåĄĒĢ┤ Ē¢źĒøä ļ¦łņØ┤Ēü¼ļĪ£ļ░öņØ┤ņś┤ņØś ņĪ░ņĀłņØä ņ£äĒĢ£ ņśłļ░® ļśÉļŖö ņ╣śļŻīņØś Ļ░ĆļŖźņä▒ņØä ņŚ┐ļ│╝ ņłś ņ׳ļŗż[33]. ņĢīņĖĀĒĢśņØ┤ļ©Ėļ│æņ▓śļ¤╝ ĒīīĒé©ņŖ©ļ│æņŚÉņä£ļÅä ņןļé┤ ļ¦łņØ┤Ēü¼ļĪ£ļ░öņØ┤ņś┤Ļ│╝ ņŚ░Ļ┤ĆļÉ£ ņŚ╝ņ”Øļ░śņØæĻ│╝ ĒśłņĢĪļćīņןļ▓Į Ēł¼Ļ│╝ļÅä ņ”ØĻ░ĆĻ░Ć ņŗĀĻ▓ĮĒć┤Ē¢ēļ│ĆĒÖöņŚÉ ņśüĒ¢źņØä ņżĆļŗżļŖö ļ│┤Ļ│ĀĻ░Ć ņ׳ļŗż[34,35]. TņäĖĒż ļ░Å TLR4 ļ¦żĻ░£ ņŚ╝ņ”Øļ░śņØæņØ┤ ņŻ╝ņÜö ĻĖ░ņĀäņ£╝ļĪ£ ņĀ£ņŗ£ļÉśņ¢┤ ņ׳ņ£╝ļéś ņČöĻ░Ć ņŚ░ĻĄ¼Ļ░Ć ĒĢäņÜöĒĢśļŗż[36,37]. ņÖĖļČĆņÖĆ ļé┤ļČĆļź╝ ļČäļ”¼ĒĢśļŖö ņןĻ│╝ ļćīņØś ņןļ▓ĮņØś ņåÉņāü(disruption of gut and blood-brain barriers)ņØĆ ņŚ╝ņ”ØņØ┤ ņל ņĀäļŗ¼ļÉśĻĖ░ ņ£äĒĢ£ ņżæņÜöĒĢ£ ĻĖ░ņĀä ņżæņŚÉ ĒĢśļéśņØ┤ļŗż[38,39].
ĻĘĖ ļ░¢ņŚÉ ņןļé┤ ļ¦łņØ┤Ēü¼ļĪ£ļ░öņØ┤ņś┤ņØ┤ ĒśĖļź┤ļ¬¼ņŚÉ ņśüĒ¢źņØä ņŻ╝ņ¢┤ ņĢīņĖĀĒĢśņØ┤ļ©Ėļ│æņŚÉ Ļ┤ĆņŚ¼ĒĢ£ļŗżļŖö ņŚ░ĻĄ¼ļÅä ņ׳ļŗż[40]. ĻĘĖļĀÉļ”░(ghrelin)ņØĆ ĒżļÅäļŗ╣ ļ░Å ņ¦Ćņ¦łļīĆņé¼ ļ┐Éļ¦ī ņĢäļŗłļØ╝ ĒĢÖņŖĄ ļ░Å ĻĖ░ņ¢Ą ļō▒ņØś Ļ│Āņ£äļćīĻĖ░ļŖź ļ░Å ļ»ĖĒåĀņĮśļō£ļ”¼ņĢä ĒśĖĒØĪņŚÉļÅä Ļ┤ĆņŚ¼ĒĢ£ļŗż. ļĀÖĒŗ┤(leptin)ņØĆ ņŗ£ņāüĒĢśļČĆņŚÉ ņ×æļÅÖĒĢśņŚ¼ Ēżļ¦īĻ░ÉņØä ļŖÉļü╝Ļ▓ī ĒĢ©ņ£╝ļĪ£ņŹ© ņŗØņÜĢņØä ņ¢ĄņĀ£ĒĢśļŖö ĻĖ░ļŖźņØä ļŗ┤ļŗ╣ĒĢ£ļŗż. ņØ┤ļ¤¼ĒĢ£ ĻĖ░ļ│Ė ĻĖ░ļŖź ņÖĖņŚÉļÅä ĻĘĖļĀÉļ”░Ļ│╝ ļĀÖĒŗ┤ņØ┤ A╬▓ņś¼ļ”¼Ļ│Āļ©ĖņŚÉ ņØśĒĢ┤ ņ£ĀļÅäļÉ£ ļÅģņä▒ņ£╝ļĪ£ļČĆĒä░ ņäĖĒżļź╝ ļ│┤ĒśĖĒĢśļŖö ņØĖņ×ÉļĪ£ ņ×æņÜ®ĒĢ£ļŗżļŖö ļ│┤Ļ│ĀĻ░Ć ņ׳ņ¢┤ ņĢīņĖĀĒĢśņØ┤ļ©Ėļ│æņØś ļ░£ļ│æĻ│╝ ņŚ░Ļ┤Ćņä▒ņØ┤ ņ׳ļŖö Ļ▓āņ£╝ļĪ£ ņāØĻ░üļÉ£ļŗż[41]. Glucagon-like peptide 1Ļ│╝ glucose-dependent insulinotropic polypeptide ņ£Āņé¼ņ▓┤ ļō▒ ĻĖĆļŻ©ņ╣┤Ļ│żĻ│╝ ņŚ░Ļ┤ĆļÉ£ ļ¼╝ņ¦łļōżļÅä ņŗĀĻ▓Įļ│┤ĒśĖĒÜ©Ļ│╝Ļ░Ć ņ׳ņ¢┤ ļ░£ļ│æņŚÉ ņØ╝ņĀĢļČĆļČä ņŚŁĒĢĀņØä ĒĢĀ Ļ▓āņ£╝ļĪ£ ņČöņĀĢļÉ£ļŗż[42,43]. ĒīīĒé©ņŖ©ļ│æņŚÉņä£ļÅä ņ£Āņé¼ĒĢ£ ĻĖ░ņĀäņ£╝ļĪ£ ļ░£ļ│æĻĖ░ņĀäņŚÉ Ļ┤ĆņŚ¼ĒĢĀ Ļ▓āņ£╝ļĪ£ ņČöņĀĢļÉśļ®░, ņØ┤ļź╝ ĒÖ£ņÜ®ĒĢ£ ņ×äņāüņŗ£ĒŚśļÅä ņ¦äĒ¢ēļÉśĻ│Ā ņ׳ļŗż[44,45].
Ļ▓░ļĪĀ
Figure 2ļŖö Ēśäņ×¼Ļ╣īņ¦Ć ņןņ£╝ļĪ£ļČĆĒä░ ļ╣äļĪ»ļÉśņ¢┤ ņŗĀĻ▓ĮĒć┤Ē¢ēņ¦łĒÖśņØś ļ░£ņāØņŚÉ ņśüĒ¢źņØä ņŻ╝ļŖö Ļ▓āņ£╝ļĪ£ ņĢīļĀżņ¦ä ņÜöņØĖļōżņØä ļÅäĒæ£ļĪ£ ņĀĢļ”¼ĒĢ£ Ļ▓āņØ┤ļŗż. ĒīīĒé©ņŖ©ļ│æĻ│╝ ņĢīņĖĀĒĢśņØ┤ļ©Ėļ│æņØĆ ļīĆļČĆļČäņØś ĻĖ░ņĀäņØä Ļ│Ąņ£ĀĒĢśņ¦Ćļ¦ī, ņןņ£╝ļĪ£ļČĆĒä░ ļ│æļ”¼ļ│ĆĒÖöĻ░Ć ņŗ£ņ×æļÉśņ¢┤ ļ»ĖņŻ╝ņŗĀĻ▓ĮņØä ĒåĄĒĢ┤ ļćīĻ╣īņ¦Ć ņĀäļŗ¼ļÉśļŖö ĻĖ░ņĀäņØĆ Ēśäņ×¼Ļ╣īņ¦ĆņØś ņŚ░ĻĄ¼ļōżņØä ļ░öĒāĢņ£╝ļĪ£ ļ│╝ ļĢī ĒīīĒé©ņŖ©ļ│æļ¦īņØś ĒŖ╣ņ¦Ģņ£╝ļĪ£ ņāØĻ░üļÉ£ļŗż. Figure 1ņØä ļ│┤ļ®┤, ŌĆśAlzheimerŌĆÖļź╝ Ļ▓Ćņāēņ¢┤ļĪ£ ņ×ģļĀźĒĢ£ Ļ▓ĮņÜ░ņØś ņ▓½ ņŚ░ĻĄ¼ļ░£Ēæ£ļŖö 1975ļģäņØĖ ļ░śļ®┤, ŌĆśParkinsonŌĆÖņ£╝ļĪ£ Ļ▓ĆņāēĒĢśņśĆņØä ļĢīļŖö 1969ļģäņØ┤ņŚłļŗż. ņ”ē, ĒīīĒé©ņŖ©ļ│æĻ│╝ ņןĻ│╝ņØś ņŚ░Ļ┤Ćņä▒ņŚÉ ļīĆĒĢ£ ņŚ░ĻĄ¼Ļ░Ć ņāüļīĆņĀüņ£╝ļĪ£ ļŹö ļ╣©ļ”¼ ņŗ£ņ×æļÉśĻ│Ā, ļŹö ļ¦ÄņØ┤ ņØ┤ļżäņ¦ĆĻ│Ā ņ׳ļŗżĻ│Ā ļ│╝ ņłś ņ׳ļŗż. ņØ┤ļŖö ņĢ×ņä£ ņé┤ĒÄ┤ļ│Ė ļ░öņÖĆ Ļ░ÖņØ┤ ĒīīĒé©ņŖ©ļ│æņŚÉņä£ ņןĻ│╝ Ļ┤ĆļĀ©ļÉ£ ļ╣äņÜ┤ļÅÖņ”ØņāüņØ┤ ļ¦ÄĻ│Ā ļ»ĖņŻ╝ņŗĀĻ▓ĮņØä ĒåĄĒĢ£ ļ│æļ”¼ļ│ĆĒÖöĻ░Ć ņל ņ”Øļ¬ģļÉśņŚłĻĖ░ ļĢīļ¼ĖņØ┤ ņĢäļŗīĻ░Ć ņČöņĀĢļÉ£ļŗż.
ņŗĀĻ▓ĮĒć┤Ē¢ēņ¦łĒÖśņØĆ ņśżļ×£ ĻĖ░Ļ░äņŚÉ Ļ▒Ėņ│Éņä£ ņä£ņä£Ē׳ ņ¦äĒ¢ēĒĢśļ»ĆļĪ£ ĒĢśļéśņØś ņøÉņØĖņØä ĒŖ╣ņĀĢĒĢśĻĖ░ ņ¢┤ļĀżņÜ┤ Ļ▓ĮņÜ░Ļ░Ć ļīĆļČĆļČäņØ┤ļŗż. ņ£ĀņĀäņĀüņØĖ Ļ░Éņłśņä▒Ļ│╝ ĒÖśĻ▓ĮņĀü ņÜöņØĖņØś ņāüĒśĖņ×æņÜ®ņ£╝ļĪ£ ņØĖĒĢ┤ ļ│æņØ┤ ļ░£ņāØĒĢśļ®░, ņØ┤ ņżæņŚÉņä£ ĻĄÉņĀĢņØ┤ Ļ░ĆļŖźĒĢ£ Ļ▓āņØĆ ĒÖśĻ▓ĮņĀü ņÜöņØĖņØ┤ļŗż. ņØ┤ļ¤¼ĒĢ£ ņĀÉņŚÉņä£ ņןļé┤ ĒÖśĻ▓ĮņØ┤ ņŗĀĻ▓ĮĒć┤Ē¢ēņ¦łĒÖś ļ░£ņāØņØś ņżæņÜöĒĢ£ ņČĢņØä ļŗ┤ļŗ╣ĒĢ£ļŗżļŖö ņĄ£ĻĘ╝ņØś ņŚ░ĻĄ¼Ļ▓░Ļ│╝ļōżņØĆ ļ»Ėņ¦ĆņØś ņäĖĻ│äņŚÉ ĒĢ£Ļ▒ĖņØī ļŗżĻ░Ćņä£Ļ▓ī ĒĢ©Ļ│╝ ļÅÖņŗ£ņŚÉ ņŗĀĻ▓ĮĒć┤Ē¢ēņ¦łĒÖś ĻĘ╣ļ│ĄņØś Ēؼļ¦ØņØä Ļ░¢Ļ▓ī ĒĢśņśĆļŗż. ĒøäņåŹ ņŚ░ĻĄ¼ļōżņØä ĒåĄĒĢ┤ ņŗĀĻ▓ĮĒć┤Ē¢ēņ¦łĒÖśņØä ļ»Ėļ”¼ ņśłņĖĪĒĢśĻ│Ā ņśłļ░®ĒĢĀ ņłś ņ׳ļŖö ļŗżņ¢æĒĢ£ ļ░®ļ▓ĢņØ┤ Ļ░£ļ░£ļÉśĻĖ░ļź╝ ĻĖ░ļīĆĒĢ£ļŗż.
 PDF Links
PDF Links PubReader
PubReader ePub Link
ePub Link Full text via DOI
Full text via DOI Download Citation
Download Citation Print
Print